In conformità con le leggi fondamentali della fisica, tutti gli esseri viventi hanno bisogno di energia dall'ambiente in qualche forma per sostenere la vita. Chiaramente, diversi organismi hanno sviluppato vari mezzi per raccogliere combustibile da varie fonti per alimentare i macchinari cellulari che guidano i processi quotidiani come la crescita, la riparazione e la riproduzione.
Ovviamente le piante e gli animali non acquistano cibo (o il suo equivalente in organismi che non possono effettivamente "mangiare" nulla) con mezzi simili, e le loro rispettive viscere non digeriscono molecole estratte da fonti di combustibile in remoto allo stesso modo. Alcuni organismi richiedono ossigeno per sopravvivere, altri ne vengono uccisi, altri ancora possono tollerarlo ma funzionano bene in sua assenza.
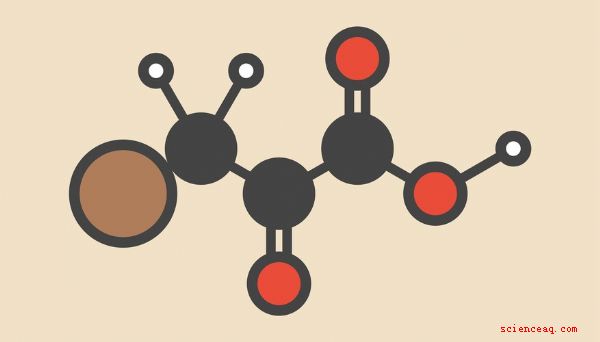
Nonostante la gamma di strategie che gli esseri viventi impiegano per estrarre energia dai legami chimici nel carbonio- composti ricchi, la serie di dieci reazioni metaboliche definite collettivamente glicolisi Una panoramica dei principali input e output della glicolisi è un buon punto di partenza per comprendere come le cellule convertono le molecole raccolte dal mondo esterno in energia per sostenere la miriade di processi vitali in cui le cellule del tuo corpo sono continuamente impegnate. I reagenti di glicolisi sono spesso elencati con glucosio e ossigeno, mentre acqua, anidride carbonica e ATP (adenosina trifosfato, la molecola vivente più comunemente usata per alimentare i processi cellulari) sono dati come prodotti per glicolisi, come segue: C 6H 12O 6 + 6 O 2 -> 6 CO 2 + 6 H 2O + 36 (o 38) ATP Chiamare questa "glicolisi", come fanno alcuni testi, non è corretto. Questa è la reazione netta della respirazione aerobica C 6H 12O 6 - > 2 C 3H 4O 3 + 2 ATP + 2 NADH + 2 H + NADH, o NAD + nel suo stato de-protonato (nicotinamide adenina dinucleotide), è un chiamato portatore di elettroni ad alta energia e intermedio in molte reazioni cellulari coinvolte nel rilascio di energia. Nota due cose qui: una è che la glicolisi da sola non è altrettanto efficace nel rilasciare ATP quanto la respirazione aerobica completa, in cui il piruvato prodotto nella glicolisi entra nel ciclo di Krebs in rotta verso quegli atomi di carbonio che atterrano nella catena di trasporto degli elettroni. Mentre la glicolisi si svolge nel citoplasma, le successive reazioni della respirazione aerobica si verificano negli organelli cellulari chiamati mitocondri. Glucosio, che contiene una struttura a sei anelli che comprende cinque atomi di carbonio e uno atomo di ossigeno, viene trasportato nella cellula attraverso la membrana plasmatica da proteine di trasporto specializzate. Una volta all'interno, viene immediatamente fosforilato, cioè un gruppo fosfato è attaccato ad esso. Questo fa due cose: dà alla molecola una carica negativa, in effetti intrappolandola all'interno della cellula (le molecole cariche non possono facilmente attraversare la membrana plasmatica) e destabilizza la molecola, dandomi più realtà suddivisa in componenti più piccoli. La nuova molecola si chiama glucosio-6-fosfato (G-6-P), poiché il gruppo fosfato è attaccato all'atomo di carbonio numero 6 del glucosio (l'unico che si trova al di fuori della struttura ad anello). L'enzima che catalizza questa reazione è una hexokinase; "hex-" è il prefisso greco per "sei" (come in "zucchero a sei atomi di carbonio") e le chinasi sono enzimi che trascinano un gruppo fosfato da una molecola e lo bloccano altrove; in questo caso, il fosfato viene prelevato dall'ATP, lasciando dietro di sé l'ADP (adenosina difosfato). Il passaggio successivo è la conversione del glucosio-6-fosfato in fruttosio-6-fosfato (F-6- P). Questo è semplicemente un riarrangiamento di atomi, o un'isomerizzazione, senza aggiunte o sottrazioni, in modo tale che uno degli atomi di carbonio all'interno dell'anello di glucosio venga spostato all'esterno dell'anello, lasciando al suo posto un anello a cinque atomi. (Si può ricordare che il fruttosio è "zucchero della frutta", un elemento dietetico comune e naturale.) L'enzima che catalizza questa reazione è la fosfoglucosio isomerasi. Il terzo passo è un'altra fosforilazione, catalizzata dalla fosfofructochinasi (PFK) e fruttosio 1,6-bisfosfato (F-1,6-BP). Qui, il secondo gruppo fosfato è unito all'atomo di carbonio che è stato estratto dall'anello nel passaggio precedente. (Suggerimento per la nomenclatura chimica: il motivo per cui questa molecola è chiamata "bisfosfato" anziché "difosfato" è che i due fosfati sono uniti a diversi atomi di carbonio, anziché essere uniti all'altro di fronte a un legame carbonio-fosfato). oltre alla precedente fase di fosforilazione, il fosfato fornito proviene da una molecola di ATP, quindi queste prime fasi di glicolisi richiedono un investimento di due ATP. La quarta fase della glicolisi rompe una molecola a sei atomi ora altamente instabile in due diverse molecole a tre atomi di carbonio: gliceraldeide 3-fosfato (GAP) e diidrossiacetone fosfato (DHAP). Aldolase è l'enzima responsabile di questa scissione. Puoi discernere dai nomi di queste molecole a tre atomi di carbonio che ognuna di esse riceve uno dei fosfati dalla molecola madre. Con il glucosio manipolato e diviso in circa uguale pezzi a causa di un piccolo apporto di energia, le restanti reazioni di glicolisi comportano il recupero dei fosfati in un modo che si traduce in un guadagno netto di energia. La ragione fondamentale per cui ciò accade è che la rimozione dei gruppi fosfatici da questi composti è più energicamente favorevole rispetto al semplice prelievo da molecole di ATP e ad applicarli ad altri scopi; pensa ai primi passi della glicolisi in termini di un vecchio adagio: "Devi spendere troppo per fare soldi." Come G-6-P e F-6-P, GAP e DHAP sono isomeri: Hanno la stessa formula molecolare, ma diverse strutture fisiche. Come accade, GAP si trova sulla via chimica diretta tra glucosio e piruvato, mentre DHAP no. Pertanto, nella quinta fase della glicolisi, un enzima chiamato triose fosfato isomerasi (TIM) prende in carico e converte DHAP in GAP. Questo enzima è descritto come uno dei più efficienti in tutto il metabolismo energetico umano, accelerando la reazione che catalizza di un fattore di circa dieci miliardi (10 10). Nella sesta fase, GAP viene convertito in 1,3-bisfosfoglicerato (1,3-BPG) sotto l'influenza dell'enzima dalla gliceraldeide 3-fosfato deidrogenasi. Gli enzimi deidrogenasi fanno esattamente ciò che suggeriscono i loro nomi: rimuovono gli atomi di idrogeno (o i protoni, se preferisci). L'idrogeno liberato da GAP trova la sua strada verso una molecola di NAD +, producendo NADH. Tieni presente che a partire da questo passaggio, ai fini contabili, tutto viene moltiplicato per due, poiché la molecola iniziale di glucosio diventa due molecole di GAP. Quindi dopo questo passaggio, due molecole di NAD + sono state ridotte a due molecole di NADH. L'inversione di fatto delle precedenti reazioni di fosforilazione della glicolisi inizia con il settimo passaggio. Qui, l'enzima fosfoglicerato chinasi rimuove un fosfato dall'1,3-BPG per produrre 3-fosfoglicerato (3-PG), con l'atterraggio del fosfato sull'ADP per formare ATP. Dal momento che, di nuovo, ciò coinvolge due molecole 1,3-BOG per ogni molecola di glucosio che entra nella glicolisi a monte, ciò significa che vengono prodotti complessivamente due ATP, annullando i due ATP investiti nei passaggi uno e tre. In fase otto, il 3-PG viene convertito in 2-fosfoglicerato (2-PG) grazie alla fosfoglicerato mutasi, che estrae il restante gruppo fosfato e lo sposta di un carbonio. Gli enzimi mutasi differiscono dalle isomerasi in quanto, anziché riorganizzare in modo significativo la struttura di un'intera molecola, spostano semplicemente un "residuo" (in questo caso, un gruppo fosfato) in una nuova posizione lasciando intatta la struttura generale. Nella fase nove, tuttavia, questa conservazione della struttura viene resa discutibile, poiché il 2-PG viene convertito in fosfoenolo piruvato (PEP) dall'enzima enolasi. Un enol è una combinazione di un alk_ene_ e di un alcool. Gli alcheni sono idrocarburi che includono un doppio legame carbonio-carbonio, mentre gli alcoli sono idrocarburi con un gruppo ossidrilico (-OH) aggiunto. Il -OH nel caso di un enol è attaccato a uno dei carboni coinvolti nel doppio legame carbonio-carbonio del PEP. Infine, nella decima e ultima fase della glicolisi, il PEP viene convertito in piruvato da l'enzima piruvato chinasi. Se sospetti dai nomi dei vari attori in questo passaggio che nel processo vengano generate altre due molecole di ATP (una per reazione effettiva), hai ragione. Il gruppo fosfato viene rimosso dal PEP e aggiunto all'ADP in agguato nelle vicinanze, producendo ATP e piruvato. Il piruvato è un chetone, nel senso che ha un carbonio non terminale (cioè uno che non si trova alla fine della molecola) coinvolto in un doppio legame con l'ossigeno e due legami singoli con altri atomi di carbonio. La formula chimica del piruvato è C 3H 4O 3, ma esprimendolo come (CH 3) CO (COOH) offre un quadro più illuminante del prodotto finale della glicolisi. La quantità totale di energia liberata (è allettante ma sbagliato dire "prodotto", poiché "produzione" di energia è un termine improprio) viene convenientemente espressa come due ATP per molecola di glucosio. Ma per essere matematicamente più precisi, si tratta anche di 88 chilojoule per mole (kJ /mol) di glucosio, pari a circa 21 chilocalorie per mole (kcal /mol). Una talpa di una sostanza è la massa di quella sostanza che contiene il numero di molecole di Avogadro o 6,02 × 10 23 molecole. La massa molecolare del glucosio è di poco superiore ai 180 grammi. Poiché, come notato in precedenza, la respirazione aerobica può derivare ben oltre 30 molecole di ATP per glucosio investito, è allettante considerare la produzione di energia della glicolisi da sola come banale , quasi senza valore. "This is completely untrue.", 3, [[Considera che i batteri, che esistono da circa tre miliardi e mezzo di anni, possono cavarsela abbastanza bene usando la sola glicolisi, perché si tratta di forme di vita squisitamente semplici che hanno pochi requisiti richiesti dagli organismi eucariotici. In effetti, è possibile vedere la respirazione aerobica in modo diverso reggendo l'intero schema sulla sua testa: mentre questo tipo di produzione di energia è certamente una meraviglia biochimica ed evolutiva, gli organismi che la usano per la maggior parte si basano assolutamente su di essa. Ciò significa che quando l'ossigeno non si trova da nessuna parte, gli organismi che si basano esclusivamente o pesantemente sul metabolismo aerobico, ovvero ogni organismo che legge questa discussione, non possono sopravvivere a lungo in assenza di ossigeno. In ogni caso, la maggior parte del piruvato prodotto nella glicolisi si sposta nella matrice mitocondriale (analoga al citoplasma di cellule intere) ed entra nel ciclo di Krebs, chiamato anche ciclo dell'acido citrico o ciclo dell'acido tricarbossilico. Questa serie di reazioni serve principalmente a generare molti portatori di elettroni ad alta energia, sia NADH che un composto correlato chiamato FADH 2, ma produce anche due ATP per molecola di glucosio originale. Queste molecole migrano quindi verso la membrana mitocondriale e partecipano alle reazioni della catena di trasporto degli elettroni che alla fine liberano 34 ATP in più. In assenza di ossigeno sufficiente (come quando ci si sta allenando intensamente), parte del piruvato subisce fermentazione, una sorta di metabolismo anaerobico in cui il piruvato viene convertito in acido lattico, generando più NAD + per l'uso nei processi metabolici.
sono comuni praticamente a tutte le cellule, sia negli organismi procariotici (che quasi tutti sono batteri) che negli organismi eucariotici (principalmente piante, animali e funghi).
Glicolisi: reagenti e prodotti
nel suo insieme, di cui la glicolisi è il passo iniziale. Come vedrai in dettaglio, i prodotti della glicolisi in sé sono in realtà piruvato e una modesta quantità di energia sotto forma di ATP:
Glicolisi: fasi iniziali
Glicolisi: fasi finali
Considerazioni energetiche e il destino del piruvato