Credito:beni comuni di Wikipedia
Una delle maggiori sfide della biologia oggi è spiegare la struttura delle creste, le membrane interne dei mitocondri. Una spiegazione in questo caso è un insieme di principi per prevedere quale forma assumeranno le creste dopo le manipolazioni metaboliche di base dell'ambiente in cui si trovano i mitocondri. Questi principi saranno quindi una descrizione della vera funzione dei mitocondri, qualcosa che finora ha solo stato appena immaginato.
I recenti progressi in tecniche come la microscopia ottica a super-risoluzione di cellule vive e la tomografia elettronica hanno fornito nuove informazioni sul comportamento dinamico delle creste. È ora possibile costruire un'architettura dettagliata dell'intero volume mitocondriale da una serie di immagini inclinate che vengono proiettate all'indietro per creare tomogrammi 3D. Lunedì abbiamo discusso di come le creste vengono trasformate e riconfigurate in base all'abbondanza e alla salute di diverse proteine della membrana interna e della matrice. Il pretesto per tale analisi erano le somiglianze strutturali e biochimiche tra le membrane dei mitocondri, dei tilacoidi e della mielina che presumibilmente aiutano a incanalare i metaboliti nella produzione di energia.
In un recente articolo di Open Biology della Royal Society , i ricercatori spiegano la biogenesi delle creste attraverso le attività coordinate di quattro percorsi principali che sono evolutivamente preservati, dai protisti e lieviti fino agli eucarioti superiori come noi:formazione di dimeri e oligomerizzazione dell'ATP sintasi ai bordi delle creste, assemblaggio del 'sito di contatto mitocondriale e sistema di organizzazione delle creste" (MICOS) alle giunzioni crista, rimodellamento della membrana da parte di una GTPasi associata alla membrana interna e correlata alla dinamina (Mgm1 nel lievito e OPA1 nei mammiferi) e corretto adeguamento della composizione lipidica della membrana.
Per il primo percorso che coinvolge l'ATP-sintasi, sono evidenti diverse cose. Come riportato in precedenza, la dimerizzazione spontanea dell'ATP-sintasi ad angoli precisi e dipendenti dalla specie in file ordinate determina la geometria del piano terra. A differenza dei complessi respiratori I-IV, che sono assemblati sulla membrana limite interna piatta, l'ATP-sintasi (complesso V) è completamente assemblato in profondità nelle membrane delle creste. Mentre diverse proteine della subunità ATP-sintasi sono superflue per una corretta formazione delle creste, le subunità Atp20 e Atp21 sono strettamente necessarie.
Un eccesso di ADP induce una conformazione condensata con ampi spazi intracristali gonfi. Al contrario, in condizioni limitanti l'ADP, i mitocondri adottano la conformazione ortodossa con spazio intracristale contratto. Nell'ameba gigante Chaos carolinensis, i mitocondri normalmente contengono creste tubolari orientate in modo casuale. Con la fame, le creste allargate adottano una morfologia cubica con un motivo a zigzag. Nei topi, gli agenti apoptotici causano la fusione delle singole creste con successivo rilascio del citocromo c dallo spazio intracristale nella regione di confine.
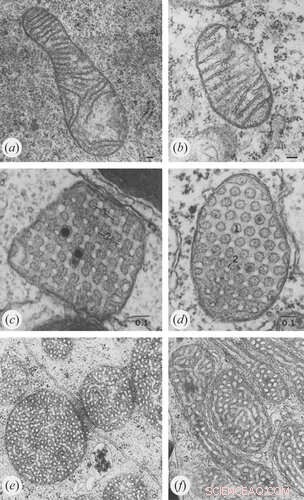
Credito:Klecker e Westermann, Biologia aperta (2021). DOI:10.1098/rsob.210238
Per il secondo percorso, l'assemblaggio dei siti di contatto MICOS, la ricerca ha determinato che proteine critiche come quelle della famiglia di geni correlati a MIC60 sono presenti fin dagli antenati endosimbiotici dei mitocondri, gli α-proteobatteri. Molti di questi precedenti precursori mitocondriali mostrano già strutture differenziate della membrana intracitoplasmatica. Invariabilmente, le specie che hanno semplificato i loro mitocondri al punto che le creste sono assenti corrispondentemente mancano dei geni correlati a MICOS. La riespressione degli omologhi MIC60 in mutanti di lievito Δmic60 salva i difetti mitocondriali ultrastrutturali.
Il terzo percorso include le GTPasi legate alla dinamina, che coordinano la fusione e la fissione delle membrane interne ed esterne. Nella fissione, queste proteine polimezzano in anelli contrattili che applicano forze costrittive per spremere i mitocondri. Il risultato è ora inteso come dipendente dalle interazioni di queste proteine, sia con il complesso MICOS che con le giunzioni creste, e anche dai sistemi di trasporto della membrana interna ed esterna che vi si riuniscono. Questi includono i complessi traslocatori di membrana TIM e TOM.
Il quarto percorso comprende gli stessi fosfolipidi della membrana mitocondriale. I mitocondri ospitano la via biosintetica della cardiolipina e sono anche coinvolti nella sintesi della fosfatidiletanolamina. Insieme alla fosfatidilcolina, questi sono i tre principali fosfolipidi con cui stanno lavorando i mitocondri. La maggior parte degli elementi costitutivi dei lipidi mitocondriali sono sintetizzati nell'ER e devono quindi essere importati da meccanismi che coinvolgono una stretta apposizione all'ER. Una volta all'interno della membrana esterna, la distribuzione dei lipidi è mediata dalle proteine di trasporto intermembrana-localizzate nello spazio della famiglia Ups/PRELI,
I mitocondri non creano la geometria da zero, ma sfruttano e si basano sulle forme fisiche naturali che si verificano spontaneamente nei lipidi. Lasciati a se stessi, i lipidi formano strutture lamellari concentriche che possono poi essere espanse e aumentate da proteine specifiche. Le misurazioni su scala fine hanno ora rivelato che le singole creste sono funzionalmente indipendenti e possono avere potenziali di membrana significativamente diversi.
La formazione delle creste implica un'interazione strettamente connessa delle quattro influenze di modellatura di cui sopra. For example, the activities of the MICOS complex and ATP synthase dimerization are both cooperative and antagonistic. MICOS induces negative membrane curvature while the ATP synthase induces positive curvature at cristae tips and rims. New computational models, as currently under development in labs throughout the world, where ratios of these different components can be finely tweaked and adjusted, will greatly aid in defining what controls the form of mitochondria.