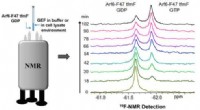
Autoinibizione: L'enzima cGAS ha un meccanismo autoinibitorio in cui domini specifici all'interno della proteina interagiscono tra loro, prevenendone l'attivazione. Questo stato conformazionale mantiene l'enzima in forma inattiva finché condizioni specifiche non ne innescano il rilascio.
Legame con proteine inibitorie: Alcune proteine, come il fattore inibitorio HERC6 (ubiquitina ligasi 6 E3 contenente domini HECT e RLD), possono legarsi al cGAS e prevenirne l'attivazione. HERC6 interagisce con l'enzima cGAS e maschera il suo sito catalitico, bloccando così la sua capacità di sintetizzare GMP-AMP ciclico (cGAMP).
Localizzazione subcellulare: Nelle cellule quiescenti, il cGAS è prevalentemente localizzato nel citoplasma, dove rimane inattivo. La presenza di DNA a doppio filamento (dsDNA) nel citoplasma, che può verificarsi durante un danno cellulare o un'infezione, innesca la traslocazione del cGAS nel nucleo, dove incontra i suoi substrati di DNA e si attiva.
Modifiche post-traduzionali: Anche le modifiche post-traduzionali, come la fosforilazione e l'ubiquitinazione, possono influenzare l'attività e la localizzazione del cGAS. La fosforilazione del cGAS da parte di alcune chinasi può potenziare la sua attività enzimatica, mentre l'ubiquitinazione può indirizzare il cGAS per la degradazione, limitandone così la disponibilità.
Questi meccanismi assicurano collettivamente che il cGAS rimanga inattivo in condizioni cellulari normali e venga attivato solo al rilevamento di stimoli specifici, come la presenza di dsDNA citosolico, che è indicativo di stress cellulare o infezione.