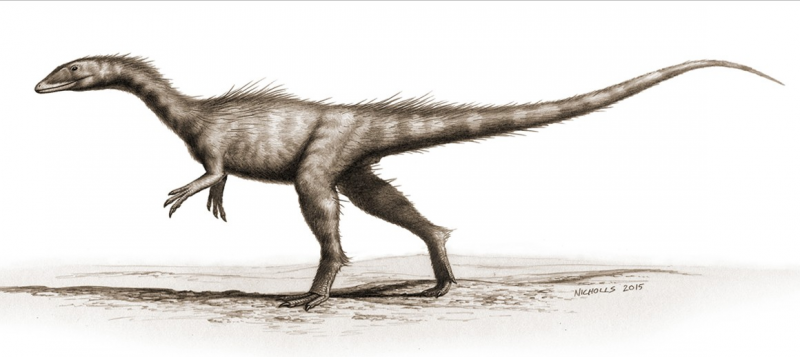
Dracoraptor hanigani (teropode) ripristinato come predatore e spazzino che abita sulla costa. Opere di Bob Nicholls (paleocreations.com). Credito:Museo di Storia Naturale, Londra. Credito:progressi scientifici, doi:10.1126/sciadv.aaw4486
L'evoluzione dell'endotermia (termoregolazione per via metabolica) rappresenta una transizione importante nella storia dei vertebrati. Però, il processo di evoluzione dell'endotermia e la sua cronologia negli uccelli e nei mammiferi rimane controverso. In un nuovo rapporto su Progressi scientifici , Enrico L. Rezende e un team di ricercatori del Centro di Ecologia Applicata e Sostenibilità, e l'Istituto di Scienze Ambientali ed Evoluzionistiche in Cile, combinato un modello di trasferimento di calore con i dati sulle dimensioni del corpo del teropode. I ricercatori hanno quindi ricostruito l'evoluzione dei tassi metabolici lungo il lignaggio dello stelo degli uccelli. I risultati hanno suggerito che una riduzione delle dimensioni costituisse il percorso di minor resistenza per l'evoluzione dell'endotermia, massimizzando l'espansione della nicchia termica, riducendo al contempo i costi di elevati fabbisogni energetici.
Di conseguenza, i ricercatori ipotizzano che il metabolismo sarebbe aumentato con la miniaturizzazione durante il periodo Giurassico Inferiore-Medio (circa 180-170 milioni di anni fa) per provocare un gradiente di livelli metabolici nella filogenesi dei teropodi. Mentre i teropodi basali possono aver mostrato tassi di metabolismo inferiori, i lignaggi non aviari più recenti erano probabilmente termoregolatori decenti con un metabolismo migliorato. L'analisi ha fornito una sequenza provvisoria nel tempo delle principali transizioni evolutive, per l'emergere di piccoli, dinosauri endotermici e piumati volanti.
L'evoluzione dell'endotermia negli uccelli e nei mammiferi è una transizione importante durante l'evoluzione dei vertebrati, fornendo un esempio straordinario di convergenza evolutiva tra gruppi, fondamentale per la loro ampia distribuzione geografica e il successo ecologico. Sebbene diversi gruppi di invertebrati e vertebrati possano aumentare le loro temperature al di sopra dell'ambiente, la capacità di mantenere alta e costante la temperatura corporea tramite la produzione di calore endogeno a riposo è esclusiva degli uccelli e dei mammiferi. L'osservazione spiegava la loro maggiore mobilità, resistenza e tolleranza in una serie di condizioni. Però, questa strategia è energeticamente costosa e non ha alcuna traccia virtuale nei reperti fossili, pertanto il ritmo e la modalità dell'endotermia rimangono controversi durante l'evoluzione dei vertebrati.
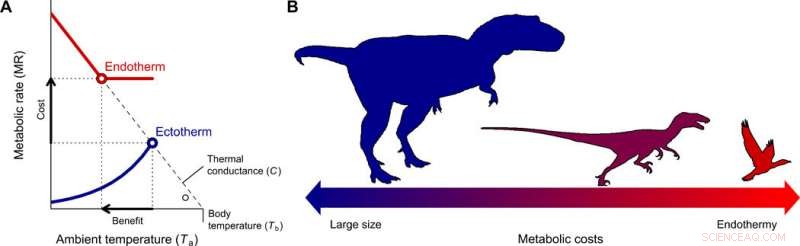
L'evoluzione dell'endotermia e la miniaturizzazione nella stirpe dei teropodi che porta agli uccelli. (A) Il costo-beneficio per passare dall'ectotermia all'endotermia per diversi intervalli di dimensioni corporee è stato quantificato con il modello Scholander-Irving, che descrive come un aumento del metabolismo a riposo (costo) aumenta la nicchia termica Tb − Ta (beneficio). Poiché non c'è gradiente termico tra l'organismo e l'ambiente in assenza di produzione di calore, questa curva interseca l'ascissa in corrispondenza di Tb =Ta quando MR =0 (8). Le linee continue blu e rosse rappresentano le curve metaboliche di un tipico ectotermo ed endoterma, rispettivamente, e i simboli aperti rappresentano il massimo gradiente termico Tb − Ta possibile con tassi metabolici a riposo, utilizzato nel nostro modello (Eq. 2). (B) Una riduzione delle dimensioni del corpo, coerente con quello descritto dai teropodi ancestrali agli uccelli basali (22), costituisce il percorso evolutivo di minor resistenza in quanto i costi energetici dell'essere grandi vengono scambiati con quelli dell'essere endotermici. Credito:progressi scientifici, doi:10.1126/sciadv.aaw4486
Per comprendere l'origine e la ragione dell'endotermia durante l'evoluzione di uccelli e mammiferi, gli scienziati hanno considerato due questioni fondamentali. Quali sono i costi ei benefici di questa strategia rispetto all'ectotermia? E quali condizioni hanno favorito una transizione verso l'endotermia? Rezende et al. affrontato le domande utilizzando il modello Scholander-Irving di trasferimento di calore, utilizzato per studiare la termoregolazione in endotermia per più di 60 anni. Il rapporto è stato usato raramente per ectotermi, ma poiché tutti gli organismi viventi producevano calore endogeno, il modello è rimasto applicabile in regime termico stazionario. Hanno fatto questo presupposto cruciale per evitare di utilizzare modelli dinamici complessi che sono spesso applicati all'ectotermia, che potrebbero rendere difficili le analisi attuali.
Hanno quantificato i costi dell'endotermia come dispendio energetico indipendente dalla massa, dove i vantaggi includevano una maggiore mobilità ed efficienza di foraggiamento, prevenzione dei predatori, tolleranza e colonizzazione di un'ampia gamma di condizioni ambientali, con aumento dei tassi di crescita e dell'omeostasi. Rezende et al. quantificato la nicchia termica che gli organismi potrebbero occupare e la sua espansione per stimare il beneficio netto dell'endotermia. Hanno quindi calcolato il costo-beneficio dell'adozione di uno stile di vita endotermico con riferimento all'antenato ectotermico e al discendente endotermico. Come originariamente proposto dal biologo evoluzionista Bran K. McNab, quando gli scienziati hanno replicato questi calcoli con stime esatte delle dimensioni del corpo, i risultati hanno mostrato che le dimensioni più piccole hanno ridotto i costi energetici per evolvere verso l'endotermia.
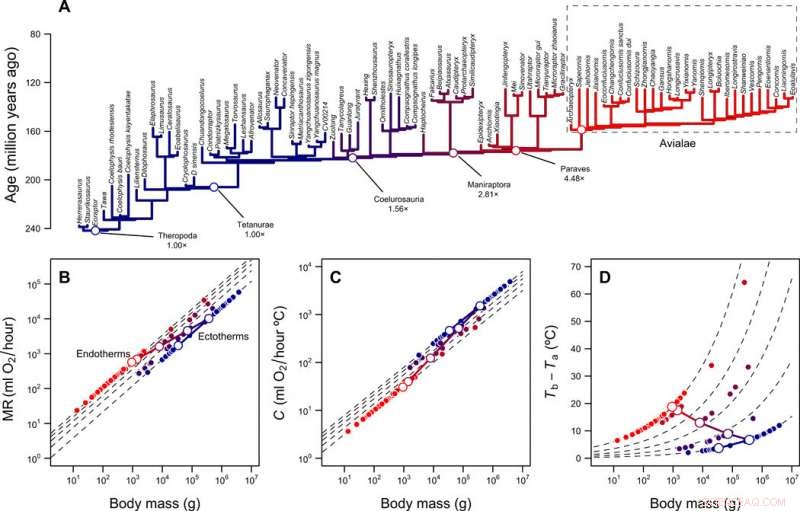
Ricostruzione dei livelli metabolici e della nicchia termica dei teropodi. (A) Filogenesi dei teropodi con rami codificati a colori in base ai livelli metabolici ricostruiti. (B) Ridimensionamento del tasso metabolico rispetto alla massa corporea per ectotermia (MR =0,68 massa 0,75) ed endotermia (MR =3,4 massa 0,75) e la traiettoria prevista del lignaggio dello stelo dell'uccello durante la transizione dall'ectotermia all'endotermia. Le linee tratteggiate mostrano le differenze di piega tra ectoterme ed endoterme (da 1× a 5×); i simboli aperti e chiusi rappresentano i valori ricostruiti per il lignaggio dello stelo dell'uccello e le punte della filogenesi, rispettivamente. (C) Ridimensionamento della conduttanza termica C e della massa corporea per ectotermi (C =2,5 massa 0,5) ed endoterme (C =1,0 massa 0,5), differenze di piega da 2,5× a 1×. (D) Gradiente termico e differenze di piega calcolate con l'Eq. 1 (nella pubblicazione) e i valori in (B) e (C). Le traiettorie lineari log-log che collegano MR e C dell'antenato ectotermico e del discendente endotermico, così come la traiettoria risultante nel gradiente termico, sono mostrati con le linee continue. Credito:progressi scientifici, doi:10.1126/sciadv.aaw4486
Il team ha esplorato come questo modello di trasferimento di calore si combinava con filogenesi e ricostruzioni delle dimensioni del corpo, gettare luce sull'evoluzione dell'endotermia negli uccelli e nei loro antenati teropodi. Rezende et al. ha stimato i costi dell'evoluzione dell'endotermia lungo il lignaggio dello stelo degli uccelli utilizzando le dimensioni del corpo ancestrali ricostruite, sulla base dei reperti fossili. Per quantificare i costi energetici all'interno di scenari alternativi hanno simulato l'evoluzione delle dimensioni del corpo lungo il lignaggio e ottenuto la distribuzione del costo per grado nel modello. Hanno assunto un modello Ornstein-Uhlenbeck (OU) non orientato tra una massa di 10 g e 100, 000 kg con un tasso evolutivo medio equivalente a quelli riportati per i teropodi. Le simulazioni hanno indicato una marcata diminuzione dei costi energetici per grado con la miniaturizzazione. Gli scienziati hanno spiegato i costi ridotti utilizzando due fenomeni.
Primo, l'espansione in nicchia termica basata su un aumento del tasso metabolico (MR) era sproporzionatamente maggiore negli ectotermi più grandi a causa della loro capacità di mantenere un'elevata temperatura corporea (T B ). Questo è stato accompagnato da un tasso metabolico indipendente dalla massa relativamente basso, dovuto all'omeotermia iniziale (termoregolazione stabile). Di conseguenza, hanno notato che maggiore è la dimensione iniziale dell'antenato ectotermico, meno costoso il passaggio all'endotermia. Secondo, durante la miniaturizzazione, gli animali hanno scambiato i costi energetici di essere grandi per essere endotermici. I risultati hanno spiegato come si sono evoluti gli alti tassi di turnover energetico, indipendentemente dal loro impatto sul fabbisogno di cibo e acqua. Nonostante la variazione intrinseca nella disponibilità delle risorse nel tempo evolutivo, in questo lignaggio sono state sistematicamente favorite dimensioni più piccole e tassi di ricambio di energia più elevati. Risultati del lavoro qualitativamente concordati con altri modelli sull'evoluzione dell'endotermia.
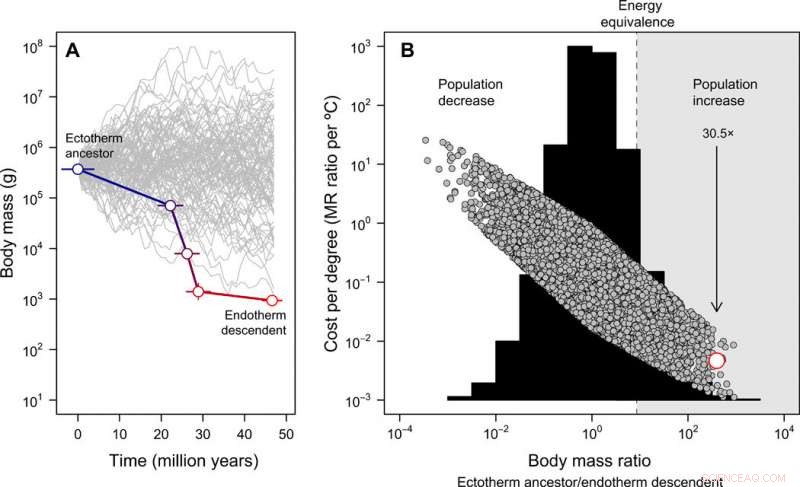
Evoluzione delle dimensioni corporee e costi-benefici dell'endotermia. (A) La miniaturizzazione da Tetanurae agli uccelli basali dedotta dai reperti fossili, contrapposto a 100 traiettorie di dimensioni simulate a partire dalla stessa dimensione corporea ancestrale a scopo illustrativo (si noti che per il successivo modello full null, le dimensioni del corpo ancestrale possono variare). L'errore rappresenta la SD in valori ricostruiti su 20 alberi candidati. (B) La distribuzione di frequenza dei rapporti di massa corporea ottenuti su 10, 000 traiettorie simulate delle dimensioni del corpo (istogramma) e i costi energetici per evolvere l'endotermia espressi per grado Celsius (Eq. 2 nella pubblicazione) sotto questo modello nullo (simboli grigi). In questo caso, la dimensione corporea ancestrale è stata ottenuta da una distribuzione uniforme compresa tra 10 g e 100, 000 chilogrammi. La stima empirica nel lignaggio dello stelo degli uccelli è mostrata in rosso. La regione in cui una riduzione delle dimensioni corporee compenserebbe i costi energetici dell'endotermia in evoluzione, consentendo alla popolazione di crescere in uno scenario di risorse costanti, è evidenziato in grigio. La freccia rappresenta l'aumento di popolazione previsto, data la riduzione delle dimensioni corporee osservata nel lignaggio dello stelo degli uccelli con l'evoluzione dell'endotermia. Queste analisi indicano che i costi energetici per evolvere l'endotermia si riducono con la miniaturizzazione e, di conseguenza, la dimensione della popolazione potrebbe essere aumentata nonostante i costi metabolici di uno stile di vita endotermico. Credito:progressi scientifici, doi:10.1126/sciadv.aaw4486
La riduzione delle dimensioni nel lignaggio dello stelo degli uccelli, strettamente abbinato al percorso teorico di minor resistenza per l'evoluzione dell'endotermia, poi Rezende et al. ricostruito come il fenomeno potrebbe essersi svolto nella filogenesi dei terapodi. Il processo ha indicato l'aumento del MR (tasso metabolico) che copre la maggior parte del Giurassico medio-alto (circa 180-170 milioni di anni fa), coinvolgendo gruppi di terapodi in cui la presenza di protopiume e piume era già onnipresente. Hanno anche suggerito tassi metabolici molto diversi tra i lignaggi concomitanti di Coelurosauria, Maniraptora e Paraves, che potrebbe spiegare l'emergere e la diversificazione di questi gruppi durante il tardo Giurassico. Il lavoro potrebbe anche spiegare la diversità anormalmente elevata di Coelurosauria a dimensioni corporee intermedie (da 30 a 300 kg) rispetto ad altri gruppi di dinosauri. Il team di ricerca ha inoltre dimostrato un modello di radiazione adattativa che riempie la nicchia nei dinosauri mesozoici con tassi eccezionali di riduzione delle dimensioni corporee nel lignaggio dello stelo degli uccelli (cioè all'interno dei nodi basali di Coelurosauria e Paraves), però, non hanno proposto un'ipotesi evolutiva adeguata per spiegare l'esito.
In questo modo, Enrico L. Rezende e colleghi hanno osservato due fenomeni eccezionali durante l'evoluzione degli uccelli; (1) una miniaturizzazione sostenuta (ma non necessariamente graduale) che abbraccia milioni di anni e (2) l'emergere dell'endotermia. Hanno usato ricostruzioni per suggerire un'evoluzione concomitante dell'endotermia con una diminuzione delle dimensioni lungo il lignaggio dello stelo dell'uccello, come già proposto in precedenza per i mammiferi. I corrispondenti cladi di terapodi hanno mostrato un intero spettro di MR. Le interpretazioni indicano che l'endotermia ha preceduto l'evoluzione del volo e la marcata riduzione delle dimensioni corporee durante l'evoluzione del lignaggio dello stelo degli uccelli è stata accompagnata da un importante cambiamento nei livelli metabolici.
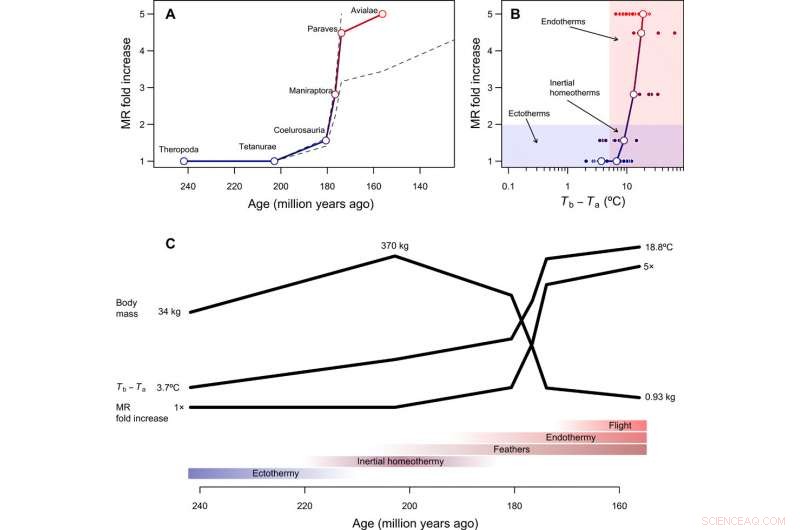
Tempo e modalità nell'evoluzione dell'endotermia. (A) Ricostruito il corso temporale dell'evoluzione metabolica nel lignaggio dello stelo degli uccelli, con linee tratteggiate che mostrano come cambiano le ricostruzioni assumendo che Paraves o Neornithes fossero completamente endotermici invece dell'uccello basale [per i calcoli con Neornithes, abbiamo ipotizzato una dimensione corporea di 150 g sulla base delle stime per Vegavis e una stima del tempo di 100 Ma fa]. L'aumento di piega della RM è stato calcolato dividendo la RM ricostruita durante la transizione all'endotermia per la RM prevista per un ectotermo di dimensioni simili ed è quindi adimensionale e indipendente dalle dimensioni del corpo. (B) Il percorso evolutivo di minor resistenza dall'ectotermia all'endotermia include l'omeotermia inerziale come fase di transizione, seguito da un aumento del metabolismo in concomitanza con una riduzione delle dimensioni. (C) Sequenza ipotetica delle transizioni evolutive nel lignaggio dello stelo degli uccelli, che combina i risultati di questo studio con ricostruzioni filogenetiche di strutture epidermiche e capacità di volo attivo (vedi il testo principale). Credito:progressi scientifici, doi:10.1126/sciadv.aaw4486
Lo scenario proposto spiegava perché i teropodi favorissero sistematicamente uno stile di vita costoso (tasso di ricambio energetico più elevato) nonostante i suoi costi energetici, e miniaturizzazione prolungata che precede l'origine degli uccelli. Per di più, uno stato di omeotermia inerziale costituiva uno stato di transizione necessario con bassi costi metabolici. Se i grandi teropodi ancestrali fossero fisiologicamente impegnati nell'omeotermia, Rezende et al. quindi si aspettano che si siano allontanati dall'allometria metabolica ectotermica (rapporto tra le dimensioni del corpo e la forma, anatomia, fisiologia e comportamento) con miniaturizzazione. Durante il periodo evolutivo, è ragionevole aspettarsi che i lignaggi sfruttino nicchie di nuova apertura e alla fine si diversifichino. I risultati di questo lavoro sono preliminari; evidenziano semplicemente che c'è probabilmente di più nell'evoluzione dell'endotermia negli arcosauri, dinosauri e uccelli di oggi. I risultati formano un'ipotesi di lavoro che in studi futuri può essere combinata con precedenti analisi sull'evoluzione delle dimensioni e di altre caratteristiche nella stirpe degli steli degli uccelli per formare interpretazioni ben definite delle sequenze temporali durante le principali transizioni evolutive.
© 2020 Scienza X Rete